本文是一篇硕士论文,硕士论文是攻读硕士学位研究生所撰写的论文。它应能反映出作者广泛而深入地掌握专业基础知识,具有独立进行科研的能力,对所研究的题目有新的独立见解,论文具有一定的深度和较好的科学价值,对本专业学术水平的提高有积极作用。(以上内容来自百度百科)今天为大家推荐一篇硕士论文,供大家参考。
前言
DNA 遗传标记的选择经历第一代遗传标记 RFLP,第二代遗传标记 STR 以及最新出现的第三代遗传标记 SNP[1]的发展过程。但由于 RFLP 有着杂合性低和分布不均匀的先天缺点很快就被第二代遗传标记即 STR 的出现所淘汰。SNP 在目前技术条件下进行大批量扩增和检测所产生的错误率较高,所需检测费用要比 STR 更高也不能幸免于淘汰。因此 STR 作为第二代遗传标记,因其独特的优点暂时无法有遗传标记可以撼动它的地位,从而得以保存。短 串 联 重 复 序 列 (short tandem repeats , STR) 又 称 之 为 微 卫 星DNA(microsatellite DNA),一种可遗传、且具高度多态性的短核苷酸重复序列[2]。由于 PCR 技术的存在,STR 片断小,序列简单因此具有容易扩增易检测的特点。由于 STR 基因座分型技术已经标准化,命名规则一致,采用完全相同的引物序列,采用不同试剂盒得出的 STR 基因座遗传信息数据结果具有可比性,因此将相同STR基因座的全部数据收集起来放在一起建立DNA数据库对数据进行共享以便日后对比查找成为一种可能。学者们将眼光投向线粒体 DNA、Y 染色体以及常染色体上多态性基因座的研究,尝试从母系遗传、父系遗传以及双亲遗传的不同角度来揭明人类的进化历史。Y 染色体和线粒体 DNA仅仅只能反映双亲中其中一条支线的演变情况,不够全面,只能作为对发生过多次重组的常染色体 STR 提供的遗传信息的重要补充[3]。通过对于不同人群的 STR 基因座多态性的研究,可以比较不同人群间的差异,计算不同群体间的遗传距离,并以此构建不同群体间的系统进化树,从而在分子生物学的水平探究不同人群的的起源、迁移、进化的过程。
国外对IdentifilerTM系统的15个STR基因座多态性调查较多,Khodjet -el-Khil[4]等对利比亚 99 个无关个体进行检测,除 CSFIPO 外,其余基因型分布均为Hardy-Weinberg 平衡,DP 和 PPE 均超过 0.999,并与北非突尼斯、摩洛哥、埃及人群的基因座相比,15 个 STR 基因座的差异均有统计学上意义。Galina petric等[5]学者对匈牙利与匈牙利在地理上和语言上邻近的人群分别进行了人群分化精确检验概率比较,得出结论伏伊伏丁那的匈牙利人与塞尔维亚人口比其他人群更具有遗传相似之处,这个可以用人口迁移和人种学杂交来解释。以上研究者均采用 IdentifilerTM系统进行检测。国内目前很多学者都有对此作了相关报道,特别是经济发达地区,已有一定的数据研究积累,1998 年俞建昆[6]等人通过 30个常染色体微卫星位点研究了 28 个东亚人群,此中就包括了汉族的南北人群,结果都支持现代中国人来源于非洲,并经由东南亚进入中国大陆。在应用于遗传学研究上,1996 年,采用五个基因频率数据,以黑猩猩的基因作为根,构建人类种群的有根系统进化树,M Nei、N Takezaki[7]这个结果证明,非洲人是第一个从上分开来的现代人类,得出结论对于关系较近的群体,使用 STR 基因座作为遗传标记进行遗传分析能提供更可靠的遗传关系。Sowcock 等通过对 14 个分布于全世界的人群的3个STR基因座进行分析,计算遗传距离并构建了系统进化树。在进化树中,先分出非洲人群,再分出欧洲人群,大洋洲人群,美洲印第安人群,亚洲人群。从中发现,其他人群应为非洲人群分化、进化的分支,进一步支持了现代人类起源的替代假说。而 2004 年 Loy.MA 等研究了委内瑞拉某地人群的遗传组成(通过对红细胞表面抗原和 STR 的标记),发现在五百年中当地美洲土著印地安人的遗传信息组成已经被不断渗入的其他人种的遗传物质冲淡。
........
第一部分 基于 IdentifilerTM检测体系对厦门汉族人群遗传多态性分析
1 材料和方法
1.1 材料
1.1.1 样本
取来自福建厦门中心血站司法鉴定所2015年3月至2016年3月的日常检案,总共 400 例无关个体,均来自厦门地区汉族人群(来本实验室要求进行亲子鉴定和个体识别的三代以上无血缘关系的厦门汉族个体)。将 PCR 扩增产物与 DNA 分子量标准 550 及去离子甲酰胺(HIDI)混匀, 95℃变性 3min,立即冷却到 4℃,将变性后的 PCR 产物在 AB 3130 型基因分析仪上进行毛细管电泳分型,选择用五色荧光运行模式(Dye Set G5)、POP-4 胶,在 36cm毛细管中电泳分型,在正负电极之间的 15KV 电压作用下 PCR 产物沿着毛细管从阴极端向阳极端迁移,由于扩增产物片段大小不同,片段的迁移率不同,扩增片段按分子量大小依次通过激光检测窗口,激光管发射激光激发 STR 基因座上标记的荧光染料产生荧光信号,利用 Data collection 软件收集荧光信号。
........
2 结果
2.1 厦门汉族人群 15 个常染色体 STR 基因座基因分型结果
15 个 STR 基因座经 X2检验获得Hardy-Weinberg平衡吻合度分析数据(表1),P>0.05,无显著的统计学差异,所以符合 Hardy-Weinberg 平衡。15 个 STR 基因座经分型得到 171 个等位基因,D3S1358、D13S317、D16S539、D18S51、D2S1338、CSF1PO、TH01、vWA、D21S11、D7S820、D5S818、TPOX、D8S1179、D19S433、FGA 15 个基因座扩增后分别发现 8、9、7、15、16、8、7、10、15、11、9、5、10、14、19 个等位基因,等位基因频率从 0.1% ~34.8%,基因频率分布如表 2 所示。
2.2 厦门汉族人群 15 个常染色体 STR 基因座相关遗传学参数
15 个 STR 遗传标记均具有高度多态性,Hets 介于 0.580~0.868, Pm 为O.036~0.148,PD 为 0.798~0.967,PIC 为 0.560~0.850,PE 为 0.268~0.730。除 CSFIPO(0.886)、D3S1358 (0.874)、TH01(0.852)、TPOX(0.798)外 DP 均大于 0.9,Hets 均大于 0.7,PIC 均大于 0.7。具体情况见表 3。
...........
第二部分 厦门汉族人群与国内其他省市人群之间的遗传关系研究......26
1 材料与方法........26
2 结果........27
3 讨论........28
4 小结........29
3 讨论
3.1 STR 基因座作为研究人群遗传关系的原因
DNA 遗传标记的选择经历第一代遗传标记 RFLP,第二代遗传标记 STR 以及最新出现的第三代遗传标记 SNP[1]的发展过程。但由于 RFLP 有着杂合性低和分布不均匀的先天缺陷很快随着时间被第二代遗传标记即 STR 的出现所淘汰。SNP 由于目前技术条件下进行大批量扩增和检测所产生的错误率较高,所需检测费用要比 STR 更高。因此 STR 作为第二代遗传标记,因其独特的优点暂时无法有遗传标记可以撼动它的地位,从而得以保存。研究者们将眼光投向线粒体 DNA、Y 染色体以及常染色体上多态性基因座的研究,尝试从母系遗传、父系遗传以及双亲遗传的不同角度来展示人类的进化历史。Y 染色体和线粒体 DNA 仅仅只能反映双亲中其中一条支线的演变情况,并不全面,只能作为对发生过多次重组的常染色体STR提供的遗传信息的重要补充[3]。通过对于不同人群的 STR 基因座多态性的研究,可以比较不同人群间的差异,计算不同群体间的遗传距离,并以此构建不同群体间的系统进化树,从而在分子生物学的水平探究不同人群的的起源、迁移、进化的过程。
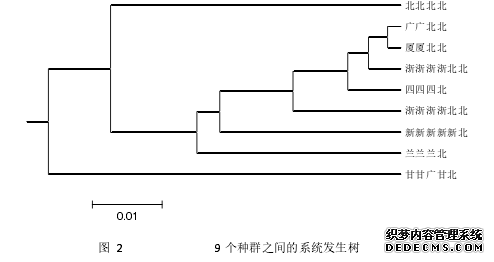
........
结论
(1) 各基因座在厦门汉族人群中具有较高的遗传多态性,基因多态性分布、基因型频率具有一定的差异,故 IdentifilerTM体系 15 个 STR 基因座具有较高法医学个人识别和亲子鉴定运用价值; IdentifilerTM检测系统的15个STR基因座并非完全适用福建厦门汉族群体,甚至中国群体的个体识别和三联体亲子鉴定的检测要求,由此,学者们有必要进一步寻求更具有地区的高鉴别力的 STR基因座。
(2) D18S51、vWA、D21S11、D7S820、FGA、TH01、D13S317、D19S433 这8 个基因座在厦门与福州、龙岩、泉州的遗传差异研究中有较大意义; 基因座D2S1338、D21S11、vWA、D7S820、D13S317 可应用于厦门与福建省外的其他群体的遗传差异研究; vWA、D21S11、D7S820、D13S317 这 4 个基因座在厦门与福建省外、省内的遗传差异性的研究中具有较大的应用价值;④各民族群体遗传背景既有特殊性和独立性,也有相互交融性。
(3)对国内 9 个群体构建系统进化树,发现福建省厦门汉族人群与广东汉族人群遗传距离较近;在不同区域、各民族之间差异较为明显。
..........
参考文献(略)

