本文是一篇医学论文,近代医学经历了16~17世纪的奠基,18世纪的系统分类,19世纪的大发展,到20世纪与现代科学技术紧密结合,发展为现代医学。(以上内容来自百度百科)今天为大家推荐一篇医学论文,供大家参考。
引 言
新城疫(Newcastle disease, ND)作为严重危害世界养禽业的一种高度传染性疾病,历来是世界各国的防治重点。NDV(Newcastle disease virus, NDV)宿主范围广、地理分布范围大,病毒在进化过程中出现了许多基因型和基因亚型,不同基因型和基因亚型NDV随着时间不断演化,在世界不同地域分布和流行,对家禽养殖业的健康发展造成严重威胁。目前普遍认为,基因VII型NDV在亚洲、欧洲、非洲以及中东等世界多个国家和地区流行,并造成了90年代以来的第四次ND大流行。随着NDV的进化,遗传差异不断扩大、进化距离逐渐变远,不断有新的基因型或亚型出现,防控形势依然严峻。目前,我国对ND实施了严格的疫苗免疫政策,La Sota株、B1株、Ulster2c株和V4株在我国以及许多其他ND流行国家应用最为广泛。然而自上世纪90年代以来,临床上非典型ND的发生变的十分普遍。许多免疫禽群即便处于较高抗体水平,但仍然时有ND的发生,进一步分析表明,这些流行株绝大部分属于基因VII型NDV。这在一定程度上表明,La Sota等传统疫苗株在抵抗VII型NDV流行株的感染中存在一定的局限性。目前普遍认为:流行株与疫苗株之间基因型和抗原性的差异是免疫禽群发生NDV强毒感染的主要原因之一,基因型匹配的新型疫苗逐渐成为研究热点。吉林大学动物医学学院传染病实验室应用基于CMV启动子的高效反向遗传操作平台拯救获得基因VII型弱毒株rmNA-1株,研究表明该毒株具有良好的疫苗种毒的前景:rmNA-1遗传稳定性良好,在鸡胚以及鸡体内连续传代仍然保持弱毒特性;安全性高,以超大剂量免疫SPF雏鸡未出现临床症状及病理变化;免疫持续期长达8周以上;与当前流行株完全匹配,可以有效减少VII型强毒攻击后的排毒及载毒。不仅如此,前期研究还表明rmNA-1具备优良的病毒活载体潜力。
目前,ND疫苗在家禽中的免疫接种几乎是必备程序。这使得NDV作为禽用疫苗载体具有良好前景。H9亚型AI(Avian influenza,AI)是严重危害养禽业的另一种传染病。目前,该病防治使用的是灭活疫苗。由于灭活疫苗在诱导黏膜免疫和细胞免疫上的局限性,无法有效抑制AIV(Avian influenza virus, AIV)在呼吸道中的复制,并且通过血清学检查不能将其与自然感染相区分,从而对AI的监测造成困难。而NDV活载体疫苗在解决以上问题方面具有巨大优势。血凝素(Hemagglutinin, HA)是诱导机体产生AI保护性抗体的主要免疫原,因此被认为是构建活载体疫苗的首选外源片段。然而,使用不同外源蛋白表达策略,HA蛋白的组装和表达势必会存在差异,进而诱导的免疫保护效果也不同。为了探究H9亚型AIV HA的最佳构建策略,本研究使用了与当前优势流行株相匹配的rmNA-1反向遗传操作平台为基础,分别构建了表达完整H9亚型AIV HA的重组病毒rmNA-H9,表达HA跨膜区(Transmembrane domain, TM)和胞质尾区(Cytoplasmic tail domain, CT)替换为NDV F基因对应区域的重组病毒rmNA-H9F,以及表达HA胞外域(Ectodomain, ECTO)并经异亮氨酸拉链(GCN4)修饰的重组病毒rmNA-H9(ECTO)。分别对3株重组病毒的生物学特性、HA蛋白表达和嵌合情况以及免疫保护效果进行了测定,以期为ND和H9亚型AI的防控提供优良的疫苗侯选株,同时为新型活载体疫苗的研究提供理论依据。
...........
第一篇 文献综述
第 1 章 NDV 分子流行病学研究进展
新城疫(Newcastle Disease, ND)是严重危害世界养禽业的一种烈性传染病,被 OIE 列为法定报告疫病,同时也是我国规定的一类动物疫病。ND 是由新城疫病毒(Newcastle Disease Virus, NDV)强毒株引起的,NDV 是不分节段的单股负链RNA 病毒,因首次分离于英格兰的新城(Newcastle upon-Tyne)以及印尼的爪哇岛(Java)而得名。
1.1 NDV 分子生物学
1.1.1 NDV 分类地位与结构基础
NDV 是副黏病毒科(Paramyxoviridae)中的禽腮腺炎病毒属(Avulavirus)的成员[1]。NDV 粒子是多形态的,但大多为球形,直径多为 100-250nm,病毒粒子被双层脂质膜包裹[2]。NDV 的基因组是含有 6 个转录单位(3'-NP-P-M-F-HN-L-5')的全长为 15186 个核苷酸、15192 个核苷酸或 15198 个核苷酸的不分段的单股负链 RNA (图 1)。基因组编码核衣壳蛋白(Nucleocapsid protein, NP)、磷蛋白(Phosphoprotein, P)、基质蛋白(Matrix protein, M)、融合蛋白(Fusion protein, F)、血凝素-神经氨酸酶蛋白(Hemagglutinin-neuraminidase, HN)和病毒 RNA 依赖性RNA 聚合酶蛋白(Large polymerase protein, L)[3, 4]。在转录过程中P基因通过“RNA编辑”机制改变读码框,从而产生两种非结构蛋白:V 蛋白和 W 蛋白[5, 6]。NDV 的 6 种结构蛋白在病毒粒子中的分布可分为两类:一类为内部蛋白,包括 NP 蛋白、P 蛋白和 L 蛋白,这三种蛋白共同参与病毒 RNA 的转录与复制[3]。另一类为外部蛋白,包括 M 蛋白、F 蛋白和 HN 蛋白。其中 M 蛋白构成囊膜内表面的支撑物。F 蛋白和 HN 蛋白是两种糖基化蛋白,以纤突的形式存在于病毒囊膜外表面,共同介导 NDV 囊膜与宿主细胞膜的吸附与融合[7]。F 蛋白是 I 型囊膜糖蛋白,其前体为无活性的 F0 蛋白,当 NDV 感染细胞后,存在于宿主细胞的蛋白酶在 F0 前体的 116 与 117 位氨基酸残基之间将其裂解为由二硫键连接的F1 和 F2 亚基,裂解后的 F 蛋白才能表现出融合活性。HN 蛋白为 II 型囊膜糖蛋白,具有血球凝集、神经氨酸酶活性,在病毒侵染过程中介导病毒粒子与细胞表面唾液酸受体的结合,并且对 F 蛋白的融合作用有促进作用。每个基因的开始和结束分别包含被称为基因启动(Gene-start, GS)和基因终止(Gene-end, GS)的控制序列。病毒 RNA 依赖 RNA 聚合酶在基因组 RNA 的 3'末端以顺序方式通过停止启动机制开始转录[8]。
......
1.2 NDV 分子流行病学
1.2.1 新城疫的流行历史与现状
目前普遍认为自1926年ND确定以来,在历史上至少发生过4次ND大流行。第一次ND的大流行发生于20世纪60年代之前,起源于东南亚地区,主要由基因II型、基因III型及基因IV型引起,持续了30多年。但由于当时养殖规模以及落后的交通运输和贸易的局限性,主要表现为局部零星暴发,而且疫情传播速度相对较慢。第二次ND的大流行主要发生于20世纪70年代,可能起源于中东地区,主要由基因V型及基因VI型引起,以基因V型为优势流行基因型。伴随国际贸易的迅速发展,此次大流行传播速度较20世纪60年代之前更加迅速。此外,野鸟及笼养鸟在ND传播过程中发挥的作用开始引发人们的注意[24-26]。第三次ND的大流行主要发生于20世纪80年代,可能起始于中东地区,主要由分离自鸽子的基因VI型NDV引发。鸽子作为传播媒介导致ND在短时间内迅速传播,而且对鸡群造成了一定的危害[27]。1985年之后,基因VII型NDV在亚洲、非洲以及中东等世界多个国家和地区流行,逐渐成为优势毒株,并造成90年代第四次ND大流行[28]。Masaji 对 2001-2007 年从日本分离的 17 株 NDV 进行分析,从鸡分离到的 7株 NDV 均为基因 VII 型[29]。Eun-Kyoun 等于 2006 年从韩国表观健康鸭分离到 1株速发型毒株,经鉴定为基因 VII 型[30]。Kang-Seuk Choi 等对 2007-2012 年从越南病死鸡中分离得到的 12 株 NDV 进行鉴定,结果均为基因 VII 型[31]。Kang-SeukChoi 于 2011-2012 年从柬埔寨病死鸡中分离到的 3 株 NDV 也全部为基因 VII 型[32]。Andrey Bogoyavlenskiy 等对 1998-2005 年从哈萨克斯坦以及吉尔吉斯斯坦分离的 28 株 NDV 进行比对分析,发现 1998-2001 年分离的 14 株 NDV 均为基因VIIb 亚型,2003-2005 年分离的 14 株均为基因 VIId 亚型[33]。TaseerAhmed Khan等对 1995-2008 年从巴基斯坦分离的 8 株 NDV 进行分析,发现其中有 4 株为基因 VII 型,另外 4 株均为基因 VI 型[34]。Mustafa M.K.Ababneh 等于 2011 年从约旦发病鸡群中分离到 1 株 NDV,经测序分析确定为基因 VII 型[35]。Sheau Wei Tan对 2004-2005 年从马来西亚分离的 8 株 NDV 进行分析,发现均为基因 VIId 亚型,与其他 NDV 分离株进行比较,发现基因 VII 型为马来西亚 NDV 主要流行基因型[36]。Sara Samadi 等对 1995-2004 年从伊朗 6 个不同地区分离的 6 株 NDV 进行分析,发现均为基因 VII 型[37]。2011-2013 年,C. Fuller 等在格鲁吉亚和保加利亚发病鸡群中分离到基因 VII 型 NDV[38]。Abdul Wajid 等于 2011-2016 年从巴基斯坦商品肉鸡、野鸡、鸽子、黑天鹅等家禽以及野鸟中分离到 52 株基因 VII 型NDV,基因 VII 型 NDV 在巴基斯坦广泛存在并持续流行,是引起巴基斯坦 ND发生流行的主要基因型[39]。2011-2016 年,Lourenc o P. Mapaco 等从非洲莫桑比克病死肉鸡中分离的 11 株 NDV 均为基因 VII 型[40]。
.........
第 3 章 表达 HA 的基因 VII 型重组 NDV 免疫特性研究.............71#p#分页标题#e#
3.1 材料............71
3.2 方法............72
3.3 结果............75
3.4 讨论............85
3.5 小结............86
第 3 章 表达 HA 的基因 VII 型重组 NDV 免疫特性研究
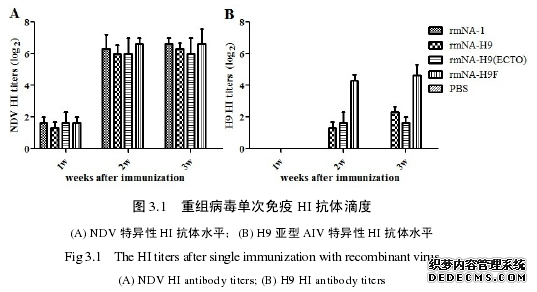
目前 H9 亚型 AI 的防控使用的均是灭活疫苗,而研究新型活病毒载体疫苗,有望提供便捷、高效的群体免疫途径,并弥补灭活疫苗在细胞免疫和局部黏膜免疫的不足。然而,以不同外源蛋白表达策略拯救获得的重组 NDV 诱导机体产生的免疫效果如何,仍有待研究。在本部分试验中,首先对 3 株重组病毒单次免疫效果进行了初步研究。将重组病毒按常规剂量通过点眼滴鼻方式免疫 7 日龄 SPF雏鸡,检测 NDV 以及 H9 亚型 AIV HI 抗体水平,并进行了动物攻毒保护试验。在此基础上,选取 rmNA-H9F 株进行两次免疫,进一步研究其免疫效力,以期为 ND 和 H9 亚型 AI 的防控提供优良疫苗侯选株,同时为重组活病毒载体疫苗的研制提供理论依据。
3.1 材料
3.1.1 病毒、SPF 鸡胚与 SPF 鸡
基因VII型NDV强毒NA-1株以及H9N2亚型AIVA/chicken/Jilin/SJ150/2012(SJ150)株由吉林大学动物医学学院传染病实验室保存。rmNA-1 弱毒株由本实验室前期拯救并保存。SPF 鸡胚购于北京梅里亚维通实验动物技术有限公司,SPF鸡使用本实验室孵化器孵化。H9 亚型 AIV 血凝抑制试验抗原、NDV 血凝抑制试验抗原购于哈尔滨维科生物;鸡新城疫、禽流感(H9 亚型)二联灭活疫苗购自乾元浩生物公司;HRP标记羊抗鸡 IgA 购自美国 BethyL 公司;可溶型单组分 TMB 底物溶液购于天根生化(北京)公司;鸡脏器淋巴细胞分离液购于天津灏洋公司;RPMI-1640 培养液购于 Hyclone 公司;流式管购自康宁公司;鼠抗鸡 CD3-FITC、鼠抗鸡 CD4-PE、鼠抗鸡 CD8a-PE、Mouse IgG1-FITC、Mouse IgG2b-PE 以及 Mouse IgG1-PE 购于美国 Southern Biotech 公司;阳离子防脱玻片购于北京鼎国昌盛公司;高品质盖玻片购于江苏世泰实验器材公司;苏木素伊红(HE)染色试剂盒购于碧云天公司。
.......
结 论
1. 基因 VII 型是近年来东北地区 NDV 优势流行基因型,并且出现新的 VIk和 VIIj 亚型。
2. 拯救了 3 株不同外源蛋白表达策略的重组 NDV,外源 AIV 血凝素蛋白的表达不影响重组 NDV 的复制,将 NDV 融合蛋白跨膜区和胞质尾区替换血凝素蛋白相应结构得到的 rmNA-H9F 株血凝素蛋白嵌合效率最高。
3. 在诱导 NDV 特异性 HI 抗体和 VII 型 NDV 攻毒保护上,3 株重组 NDV可以达到与 rmNA-1 株相似的免疫保护效果,但在诱导 H9 亚型 AIV 特异性 HI抗体和攻毒保护上,rmNA-H9F 优于另 2 株重组病毒。
4. rmNA-H9F 具有较好的免疫原性,使免疫鸡能够抵御 VII 型 NDV 和 H9N2亚型 AIV 的攻击。
..........
参考文献(略)

