本文是一篇医学论文,不管是中医学还是西医学,从二者现有的思维方式的发展趋势来看,均是走向现代系统论思维,中医药学理论与现代科学体系之间具有系统同型性,属于本质相同而描述表达方式不同的两种科学形式。(以上内容来自百度百科)今天为大家推荐一篇医学论文,供大家参考。
引 言
阴道毛滴虫(Trichomonas vaginalis, TV)呈世界性分布,是一种能够引起人类和其他动物感染的重要人兽共患寄生性原虫。它在人群中普遍感染,尤在不发达国家为甚。能够增加HIV、宫颈癌、淋病等性传播疾病的感染几率。由于阴道毛滴虫会增加HIV患病几率和不良妊娠而引起人类普遍的关注。此外,阴道毛滴虫作为真核生物分支早期的生物,是研究真核生物学特性的良好生物模型。尽管阴道毛滴虫感染被认为是一种可治愈的疾病。但是由于耐药性的增加,以及大多感染呈现隐性的特性,使得阴道毛滴虫病尚无有效的防治方法,对人类健康造成极大的危害。究其原因是对其细胞分子生物学特性研究较少。而阴道毛滴虫体外纯培养方法的建立,为阴道毛滴虫在细胞分子生物学特性上研究提供了良好的基础。众所周知,端粒是存在于真核细胞染色体末端的DNA重复序列及一些结合蛋白组成的特殊结构。保持端粒稳定,在保证细胞基因组完整、细胞长期的活性和潜在的无限增殖能力等方面具有重要作用。而在大多数真核生物中端粒的调控主要由端粒酶来实现。端粒酶是由逆转录酶(TERT,结构保守)和RNA模板(TERC,以RNA为模板,合成端粒DNA重复序列,使端粒延伸)构成。而随着端粒分子结构和功能的深入研究,端粒结合蛋白对端粒生物功能的调节作用及影响等逐渐进入人们的视野。端粒结合蛋白在维持端粒稳定,保护染色体末端完整性,端粒沉默等方面同样起着重要作用。除此外,在一些不存在端粒酶和端粒酶缺失的细胞中,存在其他的方式(如DNA重组)或DNA修复元件(Mre 11)对端粒DNA的延伸进行调节。
目前已经成功的获得了包括人类在内的大多数真核生物的端粒DNA重复序列、端粒酶中TERT序列、以及端粒结合蛋白(TBPs)序列。在原虫中,包括恶性疟原虫(TT[T/C]AGGG)、微小隐孢子虫(TTTAGG)、蓝氏贾第虫(TTAGG)、肠贾第虫(TAGGG)、利什曼原虫(TTAGGG)及布氏锥虫(TTAGGG)的端粒DNA重复序列以及端粒酶对端粒DNA调控均有研究。但阴道毛滴虫端粒DNA重复序列及其调节蛋白的研究尚未见报道。因此,本研究以阴道毛滴虫端粒DNA及其调节蛋白为研究目标,通过测序、BAL31及FISH确定阴道毛滴虫端粒DNA重复序列。对不存在端粒酶情况下,阴道毛滴虫端粒结合蛋白TRF和TBP及被称为Mre11样蛋白的SPP对端粒DNA延伸的影响,并验证PIKK与SPP相互作用和PIKK对端粒DNA延伸的影响。从而更好的为研究阴道毛滴虫端粒的调控机制及阴道毛滴虫细胞分子生物学特性等方面提供了新思路。
........
第一篇 文献综述
第一章 阴道毛滴虫研究进展
阴道毛滴虫是一种重要的人兽共患寄生性原虫,不仅能够引起人泌尿生殖系统疾病,也能够感染小鼠,使松鼠猴表现出临床症状。随着阴道毛滴虫感染对妊娠及其他疾病(如 HIV,宫颈癌等)的影响,而逐渐受到关注。阴道毛滴虫呈现典型的梨形,与宿主细胞结合时会发生形变。其滋养体长约10-20μm,宽约 2-14μm,拥有四条前鞭毛及一条延机体中部向后延伸的后鞭毛,具有波动膜[1,2]。阴道毛滴虫主要细胞外寄生于泌尿生殖道上皮以兼性厌氧的方式存活[3],碳水化合物为主要能量来源,在有氧和无氧的条件下均能够进行糖酵解[4]。具有独特的双层膜的细胞器称为氢化酶体[5]。氢化酶体能够在缺乏细胞色素,线粒体呼吸链(RC)酶和 DNA 的情况下,以不同种氢化酶体催化碳水化合物(发酵),产生的氢分子。其中只有[Fe]氢化酶,具有保守的 H-集群与四种不同的功能域。阴道毛滴虫氢化酶体包含了生物合成的所有原件,包括线粒体样 FeS 集群和公认的胞浆辅助蛋白如 FeS 蛋白[6]。阴道毛滴虫仅有滋养体阶段,所以在外界环境中并不能很好的生存,但是在潮湿的环境中能够存活三个小时以上[7]。阴道毛滴虫能够在寒冷或者其它压力条件下形成假包囊,假包囊的形式对动物具有更强的致病性,尤其对人类肿瘤发生方面相关[8,9]。此外,阴道毛滴虫能够感染 dsRNA 病毒,对其毒力及致病性均有重要意义。
........
第二章 端粒及其相关蛋白研究进展
端粒(Telomeres)是位于真核细胞染色体末端的高度重复端粒 DNA 序列和蛋白质组成的复合体,其作用是维持染色体末端的完整性和调控细胞分裂周期。端粒DNA 是由简单的 DNA 高度重复序列组成。端粒相关蛋白与端粒共同构成了特殊的“帽子”结构。端粒酶是一种由催化蛋白和 RNA 模板组成的酶,以 RNA 模板合成端粒 DNA 重复序列,用于给端粒 DNA 加尾。随着 DNA 分裂复制,端粒就会逐渐缩短,当端粒消耗殆尽时,细胞立即激活凋亡系统。为此,端粒其长度能够反映出细胞复制潜能,被称作细胞寿命的" 有丝分裂钟"。端粒是真核染色体末端特殊的核蛋白复合物。其基本结构是保守的短的富含TG 的串联重复 DNA 序列(大多数重复序列为 6-8 个核苷酸,如人 TTAGGG,贾第虫 TTAGG;但也有如酵母的不规则重复)和端粒相关蛋白形成的复合物。在染色体双链 DNA 中,富含端粒重复序列 G 的 3'端,被称为 G-链,而其互补链被称为 C-链。G-链延伸部分形成的单链称为 G-tail。在哺乳动物中,G-tail 能够折回双链端粒螺旋形成一个大型的 “T-环”,T-环的稳定性与端粒的完整性密切相关[109]。而类似的“T-环”结构在酵母、纤毛虫和锥虫中均有报道[110,111,112]。端粒也可以折叠成 G-四链体 DNA,这是基于鸟嘌呤的异常 DNA 形态[113]。
.........
第三章 端粒结合蛋白 TRF 和 TBP 对端粒 DNA 的影响....49
1 材料与方法.... 49
2 结果...... 543 讨论...... 63
4 小结...... 64
第四章 SPP 对端粒 DNA 的影响.............65
1 材料与方法.... 65
2 结果...... 72
3 讨论...... 79
4 小结...... 80
第五章 PIKK 与 SPP 相互作用及 PIKK 对端粒 DNA 的影响......81
1 材料与方法.... 81
2 结果...... 89
3 讨论...... 99
4 小结.... 100
第五章 PIKK 与 SPP 相互作用及其对端粒 DNA 的影响
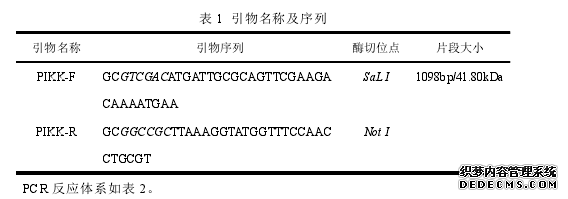
研究指出在真核生物中,3 羟基磷脂酰肌醇激酶样激酶(PIKKs)家族成员能够协调细胞 DNA 修复反应,能够结合 Mre11,迅速活化一系列下游底物磷酸化参与维持基因组的完整性[226]。而在阴道毛滴虫基因组同样发现了 PIKK 基因序列。在上一章的研究结果显示被称为 Mre 11 样蛋白的 SPP 蛋白同样对阴道毛滴虫端粒 DNA 延伸起到正调节作用。那么阴道毛滴虫 PIKK 是否能够与 SPP 结合,是否对端粒 DNA 延伸起到调节作用。本研究通过 GST pull down 及 BiFC 验证 PIKK 与 SPP 相互作用。并构建锤头状核酶,电穿孔转染至阴道毛滴虫体内,体内切割 PIKK 后,检测端粒 DNA 重复序列的变化,从而确定 PIKK 在体内对阴道毛滴虫端粒 DNA 的影响,为进一步研究端粒调节机制提供理论基础。
..........
结 论
1 首次获得阴道毛滴虫端粒 DNA 重复序列(TTTTAGGG)n。
2 阴道毛滴虫端粒结合蛋白 TRF 和 TBP 均是一种低表达的核蛋白。与阴道毛滴虫端粒 DNA 重复序列以点状形式部分共定位于细胞核染色体末端。能够在体外与阴道毛滴虫端粒双链 DNA 重复序列结合。
3 阴道毛滴虫端粒结合蛋白 TRF 和 TBP 对阴道毛滴虫端粒 DNA 延伸起到负调节作用。
4 阴道毛滴虫 SPP 蛋白能够在体外与阴道毛滴虫端粒 DNA 结合并对端粒DNA 延伸起到正调节作用。
5 阴道毛滴虫 PIKK 和 SPP 蛋白存在相互作用。且 PIKK 同样能够对阴道毛滴虫端粒 DNA 延伸起到正调节作用。
..........
参考文献(略)

