本文是一篇农学论文,农学(农业科学)是研究与农作物生产相关领域的科学,包括作物生长发育规律及其与外界环境条件的关系、病虫害防治、土壤与营养、种植制度、遗传育种等领域。(以上内容来自百度百科)今天为大家推荐一篇农学论文,供大家参考。
第一章 绪论
1.1 研究背景
氰化物是指化合物分子中含有氰根(-C≡N)的物质总称。根据与氰根连接的元素或基团的不同分成两大类,即有机氰化物和无机氰化物[1]。前者称为腈,后者常简称为氰化物。无机氰化物应用非常广泛、品种也较多,可分为简单氰化物和络合氰化物。简单氰化物包括氢氰酸(HCN),及碱金属、碱土金属和铵的化合物等,以氢氰酸和碱金属氰化物最为常见[1]。络合氰化物的种类很多,比如铁氰络合物,锌氰络合物,镍氰络合物和铜氰络合物,在环境中以铁氰络合物最为常见[2],铁氰络合物主要包括亚铁氰化物,铁氰化物。氰化物的分类如表 1-1所示。常见金属氰化络合物的稳定常数[2]见表 1-2。无机氰化物在不同环境介质间的迁移和转化与介质的理化特征和生物活动关系密切。在大气中,通常以气态的氢氰酸(HCN)为主,其主要来自于地表水中水溶态 HCN 的挥发;在地表水中氰化物的种类很多,其中以水溶态的氢氰酸(HCN)和碱金属氰化物(CN-)为主要存在形式,还有少量的铁氰化物和亚铁氰化物。简单氰化物(CN-和 HCN)在水溶液中的赋存形态很大程度上取决于水体环境的 pH,当 pH 大于 9.24,主要以 CN-的形式存在,而当 pH 小于 9.24,主导形态是水溶态的 HCN。由于铁氰化物和亚铁氰化物极易光解,它们在地表水中极为不稳定,其主要光解产物是 CN-[3,4];在土壤和地下水中, 氰化物的主要优势种类是铁氰络合物,其中铁氰化物和亚铁氰化物占到总量的 97%以上[4]。铁氰化物和亚铁氰化物在蔽光的情况下其稳定性非常好,也不会出现解离现象,但是他们在有光照或者酸性环境中(pH 小于 4.0)解离非常迅速,释放出产生剧毒的CN- [4]。由于土壤环境体系存在诸多的不确定性和复杂性,氰化物在土壤和地下水中的迁移和转化比较复杂。
..........
1.2 本课题研究目的和意义
氰化物广泛应用于合成纤维,塑料,除草剂,杀虫剂和印染业中,它还应用于贵重金属的提取,还可以作为螯合剂用于废水处理。尽管许多生物在其生化代谢的过程中能产生氰化物,但自然产生的量非常有限。而环境中的氰化物主要来源于工业活动产生的废水和废渣。2002 年美国环境保护署保守估计每年流入到环境中的氰化物大约有 10 万吨。2015 年天津港“8 12”瑞海公司危险品特别重大火灾爆炸事故,导致事故核心区水坑及事故周边区域地表水氰化物均超标。对氰化物污染的治理方法大致可分为三种:物理-化学方法、微生物降解和植物修复。物理-化学方法能快速地去除水中的氰化物,但伴随二次反应物氰酸盐的产生,而且物理-化学方法仅限于对含氰工业废水的处理。多种微生物在实验室条件下能有效地降解氰化物,但其应用于野外氰化物污染的治理尚有许多未尽事宜。大量的研究工作表明,植物不仅能快速地吸收环境中的简单氰化物,而且能有效地将其转化[7,14-16,25,26,28,49]。迄今,植物修复铁氰络合物污染的研究虽有一定的了解[17,18,27,69],但有关其修复的机理尚缺乏深入研究。本研究是在国家自然科学基金资助项目“不同代谢类型植物对铁氰化物污染修复机理的比较研究”的资助下,通过测定铁氰化物在植物体内的转移、转化和累积,比较不同代谢类型植物对其修复潜力的差异性;探讨铁氰化物做为植物生长所需补充氮源的可行性;采用基因组芯片技术分析铁氰化物胁迫下水稻幼苗的基因表达特征,为探讨植物应答氰化物胁迫的响应机制提供理论基础。
.............
第二章 不同抑制剂对植物离体根吸收和转化铁氰化物的影响
有研究显示,植物对铁氰化物的吸收可能存在两种途径[27,69]:(1)铁氰化物首先离解为三价铁离子(Fe3+)和简单氰化物(CN-),然后通过确定的运输途径进入植物的根细胞;(2)植物直接以铁氰根([FeIII(CN)6]-3)的形式吸收。对植物而言,Fe是一种必须的元素,它在植物的生物化学反应过程中发挥了重要的作用[70]。植物有多种从土壤中获取Fe的方式[71]。比如:禾本科植物可通过植物根系分泌的Fe螯合剂(麦根酸)而络合土壤中的Fe[70];一些高等植物则通过根系表面存在的Fe还原酶将三价铁转化为二价铁而被植物吸收[71]。尽管目前有一些关于植物吸收和降解铁氰化物的研究,但是其具体的吸收和代谢途径还不完全确定[7,27,28,69]。与植物吸收一些小分子的中性化学物质不同,铁氰化物不能通过简单扩散通过细胞膜[7,72]。研究显示简单氰化物可通过扩散作用快速地从溶液进入植物根细胞中[73],而且其吸收速率与外源添加抑制剂的存在与否无关[3,74]。如果在植物直接以铁氰根的形式吸收水溶液中的铁氰化物,那么外源添加的抑制剂对铁氰化物去除所造成的影响均可解释为其对铁氰化物运输过程的影响。本研究采用一个避光的全密闭反应体系来保证水溶液中铁氰化物的稳定性[27,28,69]。利用植物的离体根组织[14,16,25,48,49],在有无外源抑制剂存在的条件下,量化分析双子叶植物(柳树和大豆)与单子叶植物(玉米和水稻)对铁氰化物的吸收和转化能力。为了解不同植物离体根组织在转化铁氰化物过程中对温度变化的敏感程度,定量分析了不同植物转化铁氰化物的温度系数(Q10)。
2.1 材料与仪器
LRH—550GSI 智能人工气候箱(韶关市泰宏医疗器械有限公司);UV760CRT 双光束紫外可见分光光度计(上海傲谱分析仪器有限公司);DW-YW110A 美菱低温冰箱(中科美菱低温科技有限责任公司);电子天平 EL104(上海梅特勒-托利多仪器有限公司);R E-2000 型旋转蒸发仪(上海亚荣生化仪器厂);KQ-100E 超声波清洗器(昆山市超声波仪器有限公司);氢氧化钠、无水磷酸二氢钾、无水磷酸氢二钠、氯胺T、异烟酸、吡唑啉酮、磷酸、丙酮、乙二胺四乙酸钠、氰化钾、铁氰化钾、氯化钙、氯化汞、硝酸银、氯化镧、四乙基氯化铵、钒酸钠、2-(N-吗啡啉)乙磺酸、三羟甲基氨基甲烷(分析纯,国药集团)。垂柳(S. babylonica L.)样品来自湖南农业大学校园内; 水稻种子(O. sativaL. cv. JY 98)购买于湖南省农科院;大豆(G. max L. cv. WH)种子由湖北省农科院提供;玉米(Z. mays L. var. HK)种子由香港大学顾继东博士提供。
.........
2.2 试验方法
2.2.1 植物材料处理方法
水稻、大豆和玉米种子清洗后置于25℃的人工气候箱中采用沙培方式培育,期间定时添加25%浓度的霍格兰营养液以维持植物生长所需,幼苗培养12-15天(因植物种类不同,培养时间略有不同)后用于试验处理。柳树采用扦插的方式在室温条件下(20-25℃)培养,直至新根和叶的出现。为了去除根细胞壁上的离子,幼苗需放置于1 mM CaCl2+ 2 mM MES-TRIS缓冲溶液(pH 6.0)4小时[53]。然后将从根尖处切下6 cm左右长的根,并将其切成1 cm长的小段用于后续试验。
2.2.2 不同抑制剂对植物离体根吸收铁氰化物的影响实验
准确称取1.0 g鲜重的植物离体根组织,放置于50 mL的三角瓶中,然后,快速加入50 mL铁氰化钾溶液和抑制剂(抑制剂的浓度分别为:20 μmol/L氯化汞,50 μmol/L硝酸银,200 μmol/L氯化镧,5.0 mmol/L四乙基氯化铵,20 μmol/L钒酸钠)[53]。铁氰化钾溶液的初始浓度为10.50 (±0.225) mg CN/L。溶液的初始pH用0.1 %的氢氧化钠溶液调整到7.5。然后用锡箔纸将具塞三角瓶包裹严实,放置于人工恒温箱中,温度控制在25 ± 0.5℃,暴露时间为24小时。每次试验重复3次。
.........
第三章 铁氰化物在柳树幼苗体内的运输和代谢.............27
3.1 材料与仪器....... 28
3.2 试验方法........... 28
3.3 结果与分析....... 31
3.4 讨论..........37
3.5 本章小结........... 38
第四章 铁氰化物作为水稻幼苗生长所需补充氮源的研究...... 39
4.1 材料与仪器....... 40
4.2 试验方法........... 40
4.3 结果与分析....... 42
4.4 讨论..........50
4.5 本章小结........... 53
第五章 铁氰化物诱导水稻幼苗基因的表达分析.............54
5.1 试验材料与方法.........55
5.2 结果与分析....... 56
5.3 讨论..........79
5.4 本章小结........... 79
第五章 铁氰化物诱导水稻幼苗基因的表达分析
氰化物对生物有机体和环境都有毒害作用,而工业活动过程中产生的氰化物加速了其在环境中的负载。简单氰化物和铁氰络合物是环境中常见的氰化物[10]。而在受氰化物污染的土壤以及地下水中铁氰化物是最常见和最稳定的化合物[83]。尽管氰化物对植物有毒害作用,但在一定的浓度范围内植物可以吸收、运输和转化氰化物[3,7,27,28,78]。植物对非生物胁迫的响应非常复杂,它包括一系列的信号转导途径和转录调控等过程[120]。因此,经典的分子生物学手段己经不能满足科研的需要。基因芯片技术的发展,开启了从全基因组水平检测基因表达的时代[121]。基因芯片又称为 DNA 芯片或 DNA 微阵列,它是基于碱基互补配对的原则,将荧光或生物素标记的样品分子与 DNA 探针进行杂交,通过检测每个探针分子的杂交信号强度进而获取样品分子的数量和序列信息。常见的基因芯片包括表达谱芯片、miRNA 芯片、SNP 芯片、甲基化芯片等,可以用来检测基因表达谱、miRNA表达谱、基因突变或者进行多态性分析等,为“后基因组计划”时期基因功能的研宄提供了强有力的工具。基于高通量基因芯片技术的基因表达谱分析是当前功能性基因研究的一个重要手段,已经广泛应用于植物对逆境胁迫的研究中[122,123]。目前,许多物种的非生物胁迫研究中都采用了生物芯片的方法,例如Kreps 等使用生物芯片技术对拟南芥盐胁迫、渗透胁迫和冷胁迫 3 种逆境中的根和叶片中转录组的变化进行研究[124]。目前,在水稻上,有不少利用基因表达谱芯片进行研究的报道。郭小勤等[125]、邢俊杰等[126]研究了培矮 64S 不同处理下减数分裂期不育幼穗和可育幼穗基因表达谱的差异。Fujii 等[127]利用含有Agilent 芯片,研究了 CW 型细胞质不育系(花粉发育外形正常,在柱头上无发芽能力)与其保持系成熟花粉基因型的差异。#p#分页标题#e#
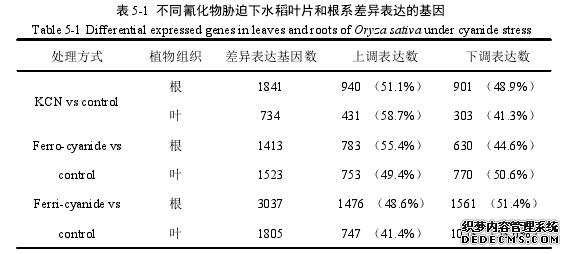
........
结论
(1)以 4 种不同代谢类型植物的离体根组织为试验材料,比较研究双子叶植物(大豆和垂柳)与单子叶植物(玉米和水稻)对铁氰化物的吸收和转化。试验结果显示:在采用的避光、封闭反应体系中,铁氰化物在反应器水溶液中发生解离的可能性很小,其在水溶液中的赋存形态是铁氰根;水稻离体根组织对反应器水溶液中铁氰化物的吸收率最快(65.8%)、其次是玉米(48.8%)、大豆最低(20.2%);而水稻离体根组织对铁氰化物的转化率也是最高的(9.11μg CN/g.h)、其次是垂柳(8.70 μg CN/g.h)、大豆离体根的转化率最低(2.79μg CN/g.h);不同的抑制剂对植物离体根组织转化铁氰化物的影响不同,氯化镧对玉米和水稻离体根转化铁氰化物的影响最大,硝酸银和四乙基氯化铵则分别对大豆和垂柳离体根转化铁氰化物的影响较大。试验温度的变化与植物离体根组织转化外源铁氰化物的效率之间存在明显的正相关性。根据对植物转化代谢铁氰化物过程的温度系数(Q10)计算发现,在 4 种不同代谢类型植物中,玉米离体根组织对温度变化最为敏感、其次是垂柳、最不敏感的是水稻。
(2)以 3 种不同种类的扦插柳树幼苗为试验材料,研究植物的蒸腾量对植物吸收铁氰化物的影响,量化铁氰化物在 3 种不同柳树种类的吸收、富集和转化的差异性。试验结果表明:在 6 天的胁迫时间内,3 种测试的柳树幼苗都可以有效吸收反应器水溶液中的铁氰化物,但吸收率在 3 种柳树幼苗间存在明显差异性,旱柳幼苗的吸收率最高(62.3%)、其次是苏柳(51.3%)、最低是垂柳(46.8%);与具有完整叶片的柳树幼苗相比,去除叶片的柳树幼苗蒸腾量呈明显的下降趋势,而且这 2 种不同状态的柳树幼苗对外源铁氰化物的吸收也存在明显的差异,表明植物的蒸腾作用对植物吸收外源铁氰化物有一定的影响;6天铁氰化物胁迫后,反应器水溶液中减少的铁氰化物只有 6.0%部分在植物体内检测到,其主要残存在柳树幼苗的根系中,其次是茎,叶片中残存量最少,这表明大部分被植物吸收到体内的铁氰化物都在体内运输的过程中都被柳树幼苗有效降解和转化,其中旱柳幼苗对铁氰化物的转化率最高(9.92 μg CN/g.d)、其次是苏柳(6.97 μg CN/g.d)、最低是垂柳(6.28 μg CN/g.d)。用水溶液和植物体内总铁浓度的变化计算铁氰化物胁迫柳树幼苗的蒸腾流浓度因子(TSCF)发现,蒸腾流浓度因子(TSCF)因柳树树种的不同而不同,其中旱柳幼苗最高(0.53)、其次是苏柳(0.48)、垂柳最小(0.3)。
..........
参考文献(略)

